テトロドトキシンとフグの耐性|なぜ自分の毒で死なないのか
テトロドトキシンとフグの耐性|なぜ自分の毒で死なないのか
フグが自分の毒で死なない理由は、ひとつではありません。テトロドトキシン(TTX、C11H17N3O8、分子量319.27)を主に外部から取り込みながら、同時に自分の電位依存性Na+チャネルのTTX結合性をアミノ酸置換で下げ、神経と筋の麻痺を避けているからです。
フグが自分の毒で死なない理由は、ひとつではありません。
テトロドトキシン(TTX、C11H17N3O8、分子量319.27)を主に外部から取り込みながら、同時に自分の電位依存性Na+チャネルのTTX結合性をアミノ酸置換で下げ、神経と筋の麻痺を避けているからです。
本稿は、「TTXはなぜ危険で、フグはなぜ平気なのか」を分子レベルで知りたい人に向けて、熱で消えない毒の基本データ、Na+チャネル外孔で何が起きるのか、Nav1.4a/bの耐性進化、臓器ごとの毒の偏り、そして研究史から2024年の検出法までをつなげて解説します。
毒性学の実務では、TTX投与前には鋭く立ち上がるNa電流トレースが、投与後にはその内向き電流だけがすっと消え、K電流の時間経過だけが残るという教科書どおりの差をまず見ます。
フグの「死なない仕組み」は、この波形の変化を頭に置くと、外因的な毒化と分子標的の耐性が噛み合った二層構造としてきれいに理解できます。
テトロドトキシンとは何か

TTXの基本データ
テトロドトキシン(Tetrodotoxin, TTX)は、フグ毒の主成分として知られる低分子毒素です。
薬理学的には電位依存性Na+チャネルを選択的に遮断する毒で、神経や筋肉の細胞膜を走る活動電位の発生と伝導を止めます。
前のセクションで触れたように、Na電流が立ち上がれなくなると、しびれから弛緩性麻痺、呼吸障害へと連続的につながります。
神経毒としての怖さは、作用点がきわめて明確で、しかも少量で深刻な生理作用を起こすところにあります。
数値を並べると、その強さが見えてきます。
化学式はC11H17N3O8、分子量は319.27です。
マウスのLD50は約8〜9 μg/kgとされ、ヒトの経口致死量は文献によって1〜2 mg、あるいは2〜3 mgと幅があります。
症状の発現は摂取後30分〜4時間の範囲に入り、食後しばらくして口唇や四肢のしびれが始まる典型像と整合します。
数字だけ見ると小さな差に見えても、mg単位のレンジで致命域に届く毒だと考えると、TTXが食中毒原因物質として特別扱いされる理由がよくわかります。
研究史の面でもTTXは独特の位置を占めます。
1909年に田原良純がフグ毒成分を単離して命名し、1964年に構造が決まり、1970年に絶対配置が確定、1972年には全合成に到達しました。
さらに1960年代以降、Na+チャネル研究の「化学的なスイッチ」として広く使われ、神経科学を押し進める道具にもなりました。
毒でありながら、イオンチャネル研究の標準試薬でもあるという二面性が、TTXの面白さです。
ミニデータボックスとして、基礎情報を一度に見渡せる形で整理すると次のようになります。
| 項目 | 内容 |
|---|---|
| 別名 | テトロドトキシン、Tetrodotoxin、TTX |
| 主な性質 | 選択的Na+チャネル遮断毒 |
| 化学式 | C11H17N3O8 |
| 分子量 | 319.27 |
| マウスLD50 | 約8〜9 μg/kg |
| ヒト経口致死量 | 1〜3 mgの範囲(1〜2 mg説、2〜3 mg説) |
| 症状発現時間 | 30分〜4時間 |
| 熱安定性 | 熱に安定、通常の加熱では失活しにくい |
熱安定性と構造の要点
TTXの実用上の厄介さは、熱に安定で、通常の加熱では失活しにくい点にあります。
つまり、食材の衛生管理でしばしば頼りにされる「火を通せば安全になる」という発想が、この毒には通用しません。
毒性学の現場感覚でいえば、TTXは“加熱調理で片づくタイプのリスク”ではなく、“最初から混入・残存させない管理”が前提になる毒です。
分子レベルでは、TTXは酸素原子を多く含む高極性の骨格をもち、グアニジニウム様の陽電荷を帯びた部位でNa+チャネル外孔近傍に強く結合します。
鍵穴にぴたりとはまるというより、チャネル入口を栓のようにふさいでNa+の流入路を止めるイメージに近いです。
これによって活動電位の立ち上がりが起こらず、感覚神経でも運動神経でも信号が伝わらなくなります。
K電流までまとめて止める非選択的な毒とは違い、Na電流に鋭く効くため、研究ツールとしても扱いやすいわけです。
⚠️ Warning
TTXの「熱に強い」と「Na+チャネルに選択的」という2点は、食中毒の危険性と研究用ツールとしての価値を同時に説明します。前者は調理で逃げ切れない理由、後者は神経科学で長く使われてきた理由です。
構造の話でもうひとつ見ておきたいのが、フグ側の耐性です。
フグのNa+チャネルにはTTXが結合できない“空白”があるのではなく、結合部位周辺のアミノ酸置換によって結合しにくくなっているのが実態です。
とくにNav1.4aとNav1.4bの系統で見つかっている置換は、TTX耐性進化の中核として扱われています。
このため、TTXそのものの構造を知ることと、受け手であるチャネル側の構造差を知ることは、表裏一体のテーマになります。
フグ以外のTTX保有生物

TTXはフグだけの専売特許ではありません。
海と陸のさまざまな生物で見つかっており、毒の分布をたどると、生合成よりも細菌起源の毒が食物連鎖を通って移るという外因的な見方が自然に浮かびます。
ここでは代表例を3つに絞って見ていきます。
まず有名なのがヒョウモンダコです。
青いリング模様の警告色で知られるこのタコは、従来は後部唾液腺にTTXを多くもつと理解されていましたが、近年は筋肉や体表にもTTXが存在し、捕食者刺激で体表粘液とともに放出されることが示されています。
咬傷では経口摂取より毒が効率よく体内に入るため、少量でも危険域に近づきます。
見た目の小ささに反して、作用点の鋭い神経毒を携えた動物です。
次にアカハライモリです。
日本で身近な両生類ですが、野生個体では高濃度のTTXや類縁体が検出されます。
一方、受精卵から無毒な餌で飼育した個体ではTTXがほとんど検出されないというデータがあり、外因性の寄与を考える材料になります。
野生幼体で1250〜13,800 ng/g、飼育群で15.2 ng/g未満という差を見ると、餌環境が毒の保有量を左右する絵がはっきりします。
皮膚の防御物質として働く、という生態的な意味づけともつながる例です。
もうひとつ挙げたいのがスベスベマンジュウガニ類、なかでもヒメスベスベマンジュウガニを含む有毒群です。
これらのカニではTTXが主要毒成分として検出された報告があり、筋肉、殻、内臓に存在すると整理されています。
ここで見落とせないのは、食用リスクが内臓だけに閉じない点です。
しかもTTXは熱に強いので、加熱調理を安全策として数えられません。
潮間帯の生物には「調理で無害化できるもの」と「そもそも食材にしてはいけないもの」がありますが、この群は後者に入ります。
TTX保有生物を眺めると、フグ、タコ、イモリ、カニと系統がばらばらです。
それでも同じ毒が繰り返し現れるのは、毒の“設計図”が生物ごとに独立して進化したというより、海洋細菌を起点にした生態学的ネットワークが背景にあるからだ、と考えると筋が通ります。
TTXは単なるフグ毒ではなく、食物網と共生微生物の境界にまたがる毒なのです。
なぜ人は麻痺し、フグは平気なのか
人での麻痺メカニズム
答えを先に置くと、人ではTTXが細胞外側から電位依存性Na+チャネルの外孔に結合し、Na+の流入を止めるため、活動電位の発生と伝導が途絶えます。
その結果、末梢神経から筋まで信号が届かなくなり、しびれ、運動麻痺、そして呼吸筋麻痺へとつながります。
フグで同じことが起きにくいのは、毒が弱いからではなく、受け手であるNavチャネル側の結合性が下がっているからです。
神経や筋肉の細胞膜では、電位依存性ナトリウムチャネルが開くとNa+が細胞内へ流れ込み、膜電位が急速に立ち上がります。
これが活動電位の出発点です。
チャネルの構造を大づかみに言えば、細胞外から見た入口に外孔があり、その少し内側にイオンを選り分ける選択性フィルターがあります。
TTXはこの入口側、つまり細胞外からアクセスできる外孔近傍に座り込み、内側へ向かうNa+の通路を物理的にふさぎます。
鍵穴の奥を壊すのではなく、玄関の外から錠前にカバーをかけて開かなくするイメージに近いです。
この作用は、電気生理の波形で見るとよくわかります。
授業や実習でよく使う典型的なトレース図では、脱分極ステップをかけた直後に鋭い内向き電流が立ち上がり、そのあと外向き成分が残ります。
TTXを加えると、まず消えるのはその最初の内向き電流です。
私はこの図を読むとき、学生には「最初の谷がNa電流、後ろに残る山やなだらかな成分が別の電流」と説明します。
TTX感受性成分を引き算で見れば、Na+チャネルがどれだけ活動電位の立ち上がりを担っているかが、一枚の図で腑に落ちます。
フグの耐性を理解する入口もここにあります。
消えるはずの成分が消えにくい、あるいは高濃度でないと消えないなら、チャネル側の薬理学が違うということです。
臨床症状とのつながりも機序どおりです。
感覚神経や運動神経の活動電位が止まれば、まず口唇や四肢の異常感覚が出て、そのあと随意運動が保てなくなります。
さらに横隔膜や肋間筋を動かす神経の伝導が落ちると、呼吸筋が麻痺します。
電線にたとえるなら、枝線が順に切れるというより、主幹スイッチが外からロックされて信号そのものが流れなくなる状態です。
筋肉そのものを壊しているわけではなく、筋に届くべき電気信号が立ち上がらないので、結果として動けなくなります。
💡 Tip
TTXは主に細胞外側からNa+チャネルの外孔に結合して遮断します。結果として活動電位の立ち上がりが抑えられるため、臨床像と分子機序の対応関係は概ね整合します(典型的な進行は感覚異常→運動麻痺→呼吸障害で、発現時間や重症度は暴露量や個体差に依存します)。
フグのNav置換によるTTX親和性低下

フグが平気な理由を厳密に言うと、「TTXの結合部位がない」ではありません。
電位依存性Na+チャネル(Nav)の外孔周辺に複数のアミノ酸置換があり、TTXとの相互作用が弱くなっているのが本質です。
つまり、同じチャネルという標的は持っているものの、分子表面の細部が変わっていて、TTXが人のチャネルほどぴたりと結合できません。
耐性の中心としてよく挙げられるのが、骨格筋型NavであるNav1.4aとNav1.4bです。
フグではこのコピーが重複しており、そこにTTX耐性と結びつく置換が見つかっています。
具体的には、ドメインIで芳香族残基が非芳香族残基に置き換わるタイプの変化や、ドメインIIでGluがAspに置換される変化が代表例です。
どちらも一文字違いのように見えて、外孔の化学的な表面性質を変えます。
芳香環の有無は立体配置や相互作用の質に響きますし、GluからAspへの置換は同じ酸性残基でも側鎖長が変わるため、TTXが結合するときの距離関係がずれます。
結果として、TTX親和性が下がります。
ここで大切なのは、フグが「絶対にTTXの影響を受けない」と単純化しないということです。
正確には、同レベルの曝露でもヒトほど機能麻痺に傾きにくいという理解が合っています。
チャネルと毒の親和性が落ちているので、外孔をふさぐ効率が下がるわけです。
毒性学では、毒の強さだけでなく、標的の感受性がどれだけ高いかで結果が決まります。
フグはまさにその標的側を変えてきた動物です。
この耐性進化は、研究者の目で見ると実に美しい設計です。
TTXは外部から蓄積する一方で、神経・筋の主要標的は生きたまま働かせなければなりません。
その矛盾を解くには、毒を隔離するだけでは足りず、標的分子の結合性も調整する必要があります。
前のセクションで触れた「外因的な毒化」と「分子標的の耐性」が二層になっているという見方は、ここで具体的な姿を取ります。
フグはTTXを抱え込みながら、Navチャネルの外孔まわりを少しずつ改造して、活動電位を維持できる側へ進化してきたのです。
TTX感受性/抵抗性Na+チャネルの比較表
哺乳類のNavチャネルにも、TTXに鋭敏なものと、相対的に抵抗性を示すものがあります。
薬理学ではこの違いをTTX-sensitiveとTTX-resistantに分けて整理します。
フグの自己耐性を理解するうえでも、この分類は見通しをよくします。
人のチャネルにももともと感受性の差があり、フグではその差をさらに耐性側へ押し進めた、と捉えると位置づけが明確になります。
| 分類 | 主なサブタイプ | TTXに対する性質 | 主な発現組織・役割 | 外孔周辺の薬理学的特徴 |
|---|---|---|---|---|
| TTX感受性 | Nav1.6 | 低濃度で阻害される | 神経軸索・ランヴィエ絞輪 | 外孔周辺がTTXと結合しやすい |
| TTX抵抗性 | Nav1.9 | 相対的に阻害されにくい | 末梢感覚神経 | 外孔周辺の残基差でTTX親和性が低い |
この表の読みどころは、TTX感受性チャネルが神経や骨格筋の活動電位に深く関わっている点です。
人でTTX中毒が起きると、しびれから運動麻痺、呼吸障害へ進むのは自然な帰結です。
末梢神経と骨格筋で活動電位が立てなくなるからです。
一方で心筋の主力であるNav1.5はTTX抵抗性に分類されます。
だからといって心臓が安全圏にあるという意味ではなく、中毒の主座がまず神経・骨格筋系に出る、という薬理学的な重心を示しています。
実習でTTX感受性電流とTTX抵抗性電流の比較図を読むとき、私は「同じNa電流でも全部が同じ顔ではない」と伝えるようにしています。
低濃度のTTXで消える成分は感受性チャネル由来、高めの条件でも残る成分は抵抗性チャネル由来という見方です。
この整理を頭に入れておくと、フグのNav置換も、単なる珍しい突然変異ではなく、外孔周辺のわずかな配列差が薬理学的性質を別物に変える実例として見えてきます。
人が麻痺し、フグが平気でいられる差は、まさにその数残基の違いに凝縮されています。
フグは毒をどう得るのか

海洋細菌起源と食物連鎖による蓄積
フグ毒をめぐる現在の理解では、TTXはフグ自身がゼロから合成するというより、海洋細菌を起点に、食物連鎖を通じて体内へ取り込まれるという外因説が主軸です。
出発点としてよく置かれるのは、海底や生物表面に生息する細菌群です。
そこから底生の無脊椎動物や小型生物に移り、さらにそれらを食べる上位の生物へと濃縮され、最終的にフグの体内へ蓄積していく、という流れです。
この話を図にするとき、私は「微生物→底生無脊椎→フグ」という三段構えをまず骨格に置きます。
そのうえで、矢印を全部同じ太さにしないほうが実態に近いと考えています。
食物網は一本の直線ではなく、途中に枝分かれや寄り道があるからです。
微生物から無脊椎への矢印は広く、無脊椎からフグへの矢印は種や食性で太さが変わる、という描き分けにすると、「どの経路が毒化へ強く寄与していそうか」が読者の目に入りやすくなります。
典型図は単純なほど伝わりますが、寄与度の差だけは矢印の太さで残しておくと、外因説の骨子が一枚で通ります。
もっとも、ここで未解明の部分も残ります。
TTXをつくる候補細菌は長く議論されてきましたが、一部の候補遺伝子や経路が示唆されているものの、遺伝子群全体の組成や機能分担はまだ確定していません。
したがって、「どの細菌が、どの条件で、どこまでTTX産生の主役なのか」は確定事項ではありません。
それでも、複数のTTX保有動物で食餌・環境依存のパターンが繰り返し見えること、そしてフグの毒化が餌環境と切り離しにくいことから、自己合成説より外因説のほうが、観察事実を無理なく説明できます。
無毒養殖フグが示す外因説の強さ
外因説の説得力を最も端的に示すのが、適切に管理された養殖環境では無毒のフグが得られるという知見です。
もしフグが体内で恒常的にTTXを自己合成しているなら、餌や飼育環境を変えても、ある程度は毒を持つはずです。
ところが実際には、毒化経路を断った養殖個体で毒が検出されない、あるいはごく低いレベルにとどまる事例が積み重なっています。
これは「毒を持つ能力」よりも、「毒を外から受け取る条件」が決定的であることを示す、強い状況証拠です。
この見方は、フグ以外のTTX保有動物でも補強されます。
たとえばアカハライモリでは、受精卵から無毒の餌で飼育した個体でTTXがほとんど検出されず、同じ地域の野生幼体では明瞭に高いレベルが出ます。
種が違っても、「野外の食物網に乗ると毒化し、切り離すと毒化しない」という方向性がそろうのは見逃せません。
フグだけを特別扱いしなくても、外因的な毒化という枠組みで整合します。
もちろん、これだけで自己合成の可能性が理論上ゼロになるわけではありません。
微量成分の処理や体内変換、保持の仕組みにはフグ自身の生理が深く関わります。
ただ、読者にいちばん伝わる論点は明快です。
フグが毒を持つかどうかは、何を食べ、どの環境に置かれたかに強く依存する。
この一点で、外因説は自己合成説より一段上の説明力を持っています。
体内動態:消化管→肝臓→皮膚/卵巣
フグがTTXを取り込んだあと、体内でどこへ運ぶかも外因説とつながっています。概略は次の流れで捉えると整理できます。
消化管 → 肝臓 → 皮膚 / 卵巣
まず食餌由来のTTXが消化管から入ります。
次に肝臓が中継点となり、その後、皮膚や卵巣といった高濃度になりやすい部位へ再分配されます。
筋肉に比べて皮膚や卵巣、肝臓で濃度が高く出やすいのは、この体内動態と生態的役割を合わせて考えると腑に落ちます。
皮膚は捕食者に対する防御の前線で、卵巣は次世代保護の場になりうるからです。
移行の細部には未解明の点が残ります。
ヒガンフグ血漿中にTTX/STX結合タンパク質(PSTBP)が報告され、輸送や保持に関与する可能性が示唆されています。
また、肝臓におけるグルタチオンやシステインとの抱合を介した一時的な処理が示唆される研究もありますが、いずれも一次報告や限定的なデータに基づく示唆段階であり、確定的なメカニズムとしてはさらに検証が必要です。
毒は体のどこに集まるのか
臓器分布
フグのTTXは、体のどこにでも均等にあるわけではありません。
分布にははっきりした偏りがあり、基本形としては卵巣と肝臓で高く、皮膚がそれに続き、筋肉は低いという並びで捉えると実態に近づきます。
前のセクションで見た「消化管→肝臓→皮膚/卵巣」という体内動態を、この臓器分布が裏づけています。
肝臓は単なる倉庫ではなく、取り込まれた毒を一時的に抱え、再配分の結節点として働く場所です。
卵巣と皮膚に高濃度が集まるのは、防御の向き先が「自分自身」と「次世代」の両方に向いていることを示しています。
組織分析でもこの傾向は明瞭です。
韓国産フグのLC-MS/MS解析では、最大1.32×10^4 μg/gという高濃度が報告されており、TTXが特定組織に強く集積することが定量的に示されています。
この種のデータを見ると、TTXは体全体に薄く散らばる毒ではなく、役割を持つ部位へ重点的に配置される毒だと理解できます。
臓器ごとの意味合いを並べると、次のようになります。
| 組織 | TTX濃度の傾向 | 生物学的な位置づけ |
|---|---|---|
| 卵巣 | 高い | 卵の保護、次世代への移行 |
| 肝臓 | 高い | 取り込み後の中継・蓄積 |
| 皮膚 | 中〜高 | 捕食者に対する最前線の防御 |
| 筋肉 | 比較的低い | 防御の主戦場ではない部位 |
ここで筋肉の扱いには少し注意が必要です。
筋肉は一般に低濃度で、卵巣・肝臓・皮膚ほどの集積部位ではありません。
ただし、だからといって「筋肉はいつでも同じ」「どの種でも一律に低い」と単純化すると、分布の実像を取りこぼします。
TTXの偏りは種の違い、成熟状態、季節要因と結びついて動くため、筋肉は相対的に低い部位として位置づけるのが適切です。
ここでも大事なのは、低い部位があることより、高い部位がなぜそこなのかです。
卵巣・皮膚・肝臓に毒が集まる配置には、生理と生態の両方の理由があります。
雌雄差・発生段階・季節性
TTX分布は固定された地図ではなく、繁殖周期に合わせて動きます。
とくに目立つのが雌の卵巣で濃度が高まりやすいという傾向です。
これは単なる雌雄差ではなく、産卵というライフイベントに向けて毒の置き場所が調整されている、と見るほうが自然です。
卵巣に高濃度のTTXが集まるなら、その毒は雌の体を守るためだけでなく、卵へ受け渡される防御資源としても読めます。
季節性もこの解釈とよく噛み合います。
産卵期に近づくにつれて卵巣中のTTXが目立ち、皮膚分布とのバランスが変化するパターンは、繁殖防御を優先する体内再配置として理解できます。
皮膚は外敵に対する前線、卵巣は次世代の防護壁です。
フグは一年を通して同じ場所に同じ濃度で毒を置くのではなく、「いま守るべき対象」に合わせて配分を変える生物だと考えると、この季節変動は筋が通ります。
概念図にすると、季節変動は次のように整理できます。
| 時期 | 卵巣のTTX | 皮膚のTTX | 読み取れる生態的意味 |
|---|---|---|---|
| 非繁殖期 | 中 | 中〜高 | 自身の体表防御が主軸 |
| 産卵期前後 | 高 | 中〜高 | 卵保護への重点移行 |
| 発生初期 | 卵・仔魚に移行 | — | 次世代防御の立ち上がり |
発生段階まで視野を広げると、TTXは親個体の属性ではなく、生活史全体をまたぐ防御因子として見えてきます。
卵の段階で毒を持つことには意味があります。
遊泳能力が乏しい、逃げられない、反撃できないという初期発生段階では、行動より化学防御の比重が上がるからです。
成魚では皮膚と行動、卵では化学的な不味さや毒性そのもの、と防御の形が切り替わるわけです。
卵と仔魚の防御:生態学的意義
卵や仔魚にTTXがあることの意味は明快です。
食べられやすい段階を、先回りして守るためです。
産み落とされた卵はその場を動けず、仔魚も発生初期には遊泳能力が限られます。
こうした時期に毒を持たせるのは、鎧を着せるというより、「ひと口で学習させる標識」を付けるのに近い発想です。
捕食者にとって不快な経験が一度でも成立すれば、その後の回避行動につながります。
ここで注目したいのは、TTXが単なる致死毒ではなく、警告シグナルと結びつく防御資源として働く点です。
化学的には食べたときの不利益を与え、行動学的には「この餌は危ない」という学習を誘導する。
皮膚に毒が集まる成魚では接触・摂食の場面で防御が成立し、卵や仔魚では小さな体を補う化学的バリアになります。
毒の配置を生態の文脈に戻すと、卵巣に高濃度、皮膚に次点、筋肉に低濃度という分布が、ただの分析値の差ではなく、捕食圧に対して最適化された配置として読めます。
この視点は、フグ以外のTTX保有動物を見るとさらに納得しやすくなります。
たとえばアカハライモリでは皮膚表面の防御、ヒョウモンダコでは体表粘液とともに放出される防御が確認されており、TTXは「体内にある毒」で終わらず、「外敵と接する面に置かれる毒」として機能します。
フグの卵や仔魚も同じ延長線上にあります。
動けない段階ほど、化学防御は前面に出ます。
💡 Tip
臓器分布を見ると、TTXは「たまたま溜まった毒」ではなく、「皮膚で自分を守り、卵巣で次世代を守るよう配分された毒」と捉えるほうが実態に合います。
筋肉の低濃度も、この生態学的配置の裏返しです。
防御の効率だけを考えるなら、捕食者が最初に触れたり口にしたりする場所、あるいは卵へ移せる場所に置くほうが合理的です。
筋肉は運動のための組織であって、防御の最前線ではありません。
だからこそ、臓器分布を読むときは「どこに多いか」だけでなく、「なぜそこに置くと有利なのか」まで踏み込むと、フグ毒の役割が一段はっきり見えてきます。
フグ毒研究は何を変えたのか
1909/1964/1970/1972:解明の節目
フグ毒研究の転機は、危険な天然毒を「正体不明の脅威」から「扱える分子」へ変えていった点にあります。
起点として押さえたいのが1909年です。
田原良純がフグ毒の有効成分を単離し、テトロドトキシンと命名しました。
この時点では、毒性は知られていても、分子として何者なのかはまだ霧の中にありました。
にもかかわらず、単離と命名が行われたことで、フグ毒は民間知識や食中毒の経験則ではなく、化学と薬理の対象として研究室へ持ち込まれます。
その後の1960年代に、TTXは電位依存性Na+チャネルをふさぐことで興奮性を止める、という仮説が前面に出てきます。
ここが研究史の要です。
神経と筋肉の活動電位はNa+の流入で立ち上がりますが、その入口だけを選んで止められるなら、膜興奮の仕組みを分解して調べられます。
TTXは単なる猛毒ではなく、神経の配線図を読むための“切り分けナイフ”になりうる、と見えてきたわけです。
1964年には3つの研究グループが相次いで構造決定を発表し、TTX研究は一気に加速します。
化学構造が定まると、どの官能基が活性に効くのか、なぜ水溶性が高いのか、なぜNa+チャネル外孔と強く相互作用するのかという議論が、推測ではなく分子の形に基づいて進められるようになります。
毒性学と神経科学が、ここで同じ分子を共有し始めました。
1970年にはX線結晶構造解析によって絶対配置が決まり、立体化学の曖昧さが消えます。
天然物化学では、この「左右どちらの立体なのか」が詰まらないと、全合成も作用機序の精密理解も止まります。
さらに1972年、岸義人らがラセミ体の全合成に到達しました。
天然由来の希少物質だったTTXが、化学的に再現可能な対象へ移った瞬間です。
毒の研究はここで一段変質し、採れるかどうかに左右される観察科学から、構造・活性相関を組み立てる設計科学へ踏み込みました。
神経科学の道具としてのTTX
TTXが神経科学に残した最大の遺産は、chemical toolとしての完成度の高さです。
膜電位の変化にはNa+、K+、Ca2+など複数のイオン電流が絡みますが、TTXを入れるとNa電流だけを選択的に切り分けられます。
すると、活動電位の立ち上がりにNa+流入がどこまで必要か、残ったK電流やCa2+電流がどのように波形を作るかが見えるようになります。
電気生理学の実験でTTXが一種の標準試薬として扱われてきた理由はここにあります。
この選択性は、神経細胞を相手にした実験でとくに威力を発揮します。
たとえばニューロンの発火が止まったとき、それが細胞死なのか、Na+チャネル遮断による可逆的な沈黙なのかを切り分けるには、作用点が明瞭な化合物が必要です。
TTXはその条件を満たしていました。
実験系に加えると、活動電位依存のシグナルだけが静まり、受容体、シナプス後応答、Ca2+依存過程など別の要素を順番に観察できます。
神経回路研究で「この反応はスパイク依存か」を問う場面にTTXが頻出するのは、測りたい現象を一段ずつ剥がせるからです。
病態研究でもTTXの役割は大きく、Na+チャネル機能異常が関わるチャネル病の理解を支えてきました。
骨格筋の過興奮、不整脈、てんかん、疼痛の一部では、Na+チャネルの開き方・閉じ方のズレが症状の核になります。
TTXはそこに直接作用するため、「Na+チャネル由来の異常かどうか」を検証する基準線として使えます。
薬理学の言葉でいえば、TTXは治療薬そのものというより、病態を見分けるための輪郭線を引く分子です。
脳科学辞典:フグ毒や公的・学術的なまとめ(例:PubChem の Tetrodotoxin エントリ)で整理されている研究史を読むと、TTXの価値は「毒が強い」ことではなく、「狙う場所が明確で余計なノイズを増やさない」点にあるとわかります。
。
ℹ️ Note
TTXの本質は、神経活動を乱雑に壊す毒ではなく、Na+チャネルという一点をふさいで電気信号を止める“分子プローブ”であるということです。
TTX-s/TTX-r研究がもたらした視界
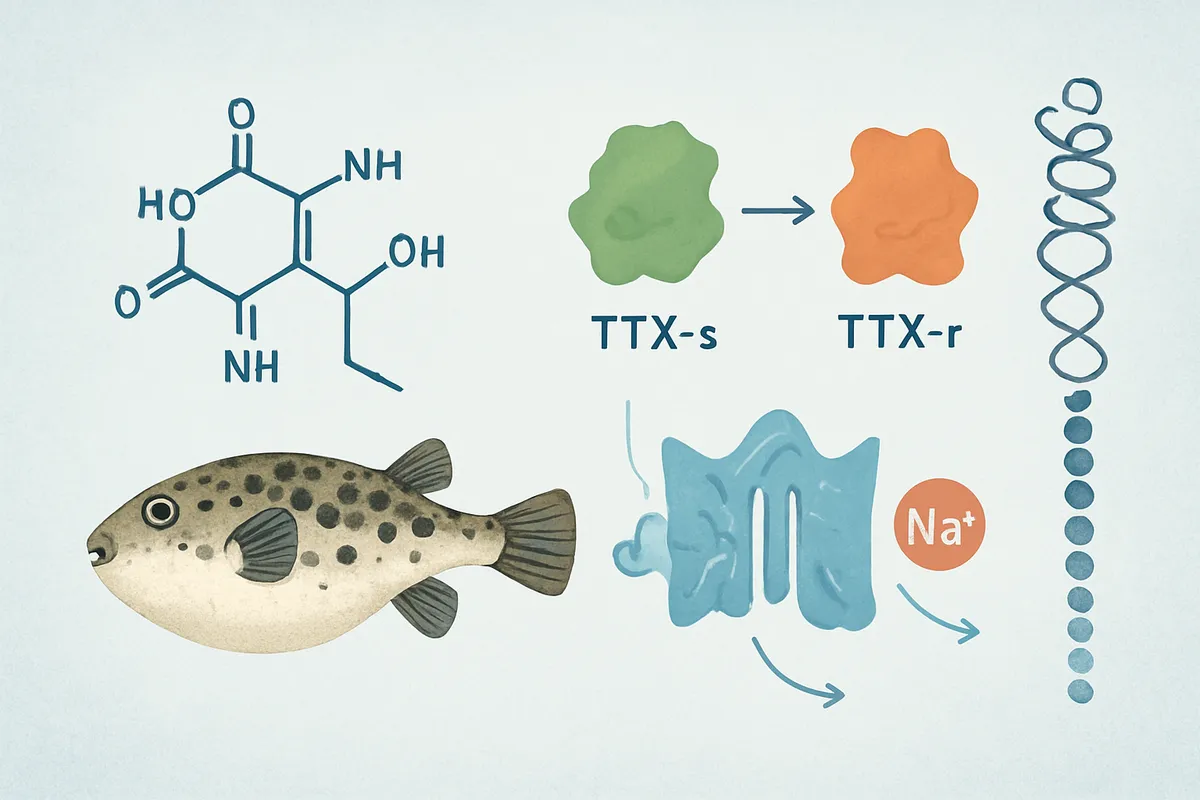
TTX研究がもう一段深い意味を持ったのは、Na+チャネルを「TTXで止まる群」と「止まりにくい群」に分ける視点を生んだからです。
ここで使われるのがTTX-s(tetrodotoxin-sensitive)とTTX-r(tetrodotoxin-resistant)という分類です。
前者は低濃度のTTXで抑えられるチャネル、後者は抑えられにくいチャネルで、同じNa+チャネルでも薬理学的な顔つきが違うことが見えてきました。
この分類は、単なる感受性のラベルではありません。
神経系のどこに、どんな役割のチャネルがいるのかを読み解く入口になりました。
中枢ニューロンや多くの軸索で主役になるチャネルはTTX-s側に寄り、痛覚や一部の末梢神経、心臓近傍の特殊な系ではTTX-r側の性質が意味を持ちます。
つまりTTXを使うと、「Na+チャネルがあるか」だけでなく、「どの型のNa+チャネルが働いているか」まで薬理学的に仕分けられるのです。
この仕分けは、その後の分子遺伝学へ自然につながりました。
のちにNav1.1、Nav1.2、Nav1.4、Nav1.7といったサブタイプ多様性が明らかになると、TTX-s/TTX-rという古典的な薬理分類が、分子実体を持った地図として読み直されます。
先に薬理で輪郭を描き、後から遺伝子とタンパク質がその輪郭を埋めた形です。
神経科学では、こうした「薬で見えた差が、のちに分子の差として確定する」流れが強い発見になります。
TTX研究はその好例でした。
フグ自身の耐性研究とも、ここはきれいにつながります。
フグのNa+チャネルは外孔周辺のアミノ酸置換によってTTXが結合しにくくなっており、毒を持つ側が同じ標的をどう回避するのかが分子レベルで読めます。
研究者の目線で見ると、TTX-s/TTX-rの枠組みは、ヒトの神経生理、疼痛研究、チャネル病、そしてフグの自己耐性までを一つの座標系に載せた、と言えます。
フグ毒研究が変えたのは「毒の知識」だけではありません。
Na+チャネルという巨大な研究分野そのものの見え方でした。
最新研究:検出法と合成研究の現在地
HILIC-LC-MS/MS:研究・公的検査の基盤
TTXの分析で今も中核にあるのは、HILIC-LC-MS/MSを軸にした高精度定量です。
極性が高いTTXは逆相系だけでは扱いにくく、親水性相互作用クロマトグラフィーを組み合わせた設計がよく噛み合います。
研究現場でも公的検査でもこの方式が重宝されるのは、単に「よく測れる」からではありません。
TTX本体だけでなく、類縁体を含む多成分解析に展開しやすいからです。
食品衛生、野生生物の毒性分布、組織別の蓄積研究を同じ分析思想の延長でつなげられる点が強いのです。
この強みは、2023年以降の組織分析データを読むとよく見えます。
たとえば韓国産フグの組織分析では、LC-MS/MSベースの定量により最大 1.32×10^4 μg/gという高濃度例まで拾われています。
卵巣や肝臓で高く、筋肉で相対的に低いという既知の傾向を、種差や個体差を含めてきちんと比較できるのは、こうした定量法の土台があるからです。
アカハライモリの飼育群と野生群の差をみた研究でも、HILIC-LC-MSが外因性起源の議論を押し進めました。
毒の「ある・ない」ではなく、「どこに、どれだけ、どの成分がいるか」を一段深く読めるのがこの系統の価値です。
2024年の検出法レビューでも、TTX分析の基盤としてLC-MS/MS系が整理されています。
現場向けの新技術が増えても、確認検査、精密定量、法的・公的判断に近い用途ではLC-MS/MSが基準線です。
感度競争だけなら別の手法が前に出る場面もありますが、マトリクスの複雑な試料を相手にして再現性と特異性を両立するとなると、結局ここへ戻ってきます。
毒性学の目線で言えば、LC-MS/MSは「見つける装置」というより、議論を確定させる装置です。
スマホ蛍光・PEC・自動パッチ:迅速・高感度の最前線

ここ数年で鮮度が上がったのは、“分析室の外”を意識したTTX検出です。
2024年には、スマホ型蛍光バイオセンサーが、測定時間約15分という現場志向の仕様で報告されました。
スマートフォンを読み取り系に組み込む設計は、装置全体を小さくまとめやすく、一次スクリーニングとの相性がよいのが特徴です。
公的検査の代替というより、「持ち帰って精密分析する前に、まず危険信号を拾う」という役回りがはっきりしています。
研究室に大型機器がなくても入口の判断を作れる、という意味で応用範囲は広いです。
同じ2024年には、近赤外光応答PECマルチチャネル免疫センサーも登場しています。
こちらは迅速性だけでなく、TTXのLODが7 pg/mLという感度が目を引きます。
さらにOAも同時に扱えるマルチチャネル設計なので、海産毒の複合リスクを一つの測定系で見る発想に進んでいます。
LC-MS/MSが「精密で幅広い」なら、PECや免疫センサーは「狙った標的を鋭く追う」方向です。
試料前処理、装置運用、解析の重さまで含めて考えると、これは競合というより役割分担の細分化です。
もうひとつ見逃せないのが、自動パッチクランプ検出です。
TTXはNa+チャネル遮断毒なので、濃度そのものを化学分析で測る方法とは別に、チャネル阻害という機能で捉えることができます。
自動パッチクランプは、従来の手作業パッチクランプより測定の回転が上がり、チャネル電流がどの程度落ちるかを短時間で評価できます。
これは食品検査の一次判定というより、薬理学的活性の確認、類縁体の機能差、チャネルサブタイプごとの応答比較に強い流れです。
TTX研究が神経科学の道具として発展してきた歴史を思えば、検出法が再び“機能測定”へ戻ってきたのは自然な展開でもあります。
手法ごとの向き不向きを整理すると、現在地は次のようになります。
| 手法 | 主な強み | 向く用途 | 現在の位置づけ |
|---|---|---|---|
| LC-MS/MS(HILIC-LC-MS/MSを含む) | 高精度・多成分解析 | 研究、公的検査、確認定量 | 基盤技術 |
| 免疫・PECセンサー | 高感度 | 標的毒の迅速高感度測定 | 専用用途で拡大中 |
| スマホ型蛍光バイオセンサー | 迅速・現場適用 | 現場スクリーニング、簡易判定 | 実装志向の新潮流 |
| 自動パッチクランプ | 薬理機能の直接評価 | チャネル阻害評価、研究 | 機能的検出の前線 |
💡 Tip
TTXの検出は、濃度を測る化学分析と、Na+チャネルをどれだけ止めるかを見る機能評価の二本立てで進んでいます。前者は「何がどれだけあるか」、後者は「それが生理機能にどう効くか」を切り分ける発想です。
合成化学と生合成模倣:現在地と課題
TTXの合成研究は、1972年のラセミ体全合成以後、多段階合成をどう整理し、どう再現性と供給性を上げるかという方向で積み重なってきました。
TTXは官能基が多く、立体配置の制御も厳しく、天然物合成のなかでも「難物」の部類です。
そのため合成研究の価値は、単に作れたかどうかでは決まりません。
どの段階で立体を組み上げ、どこで保護基戦略を簡素化し、最終的に類縁体研究へつなげられるかまで含めて評価されます。
2025年から2026年にかけての総説では、この流れが「歴史の回顧」ではなく、供給・誘導体展開・生合成理解を束ねる課題として整理されています。
研究の重心は、純粋な全合成の洗練だけではありません。
近年は生合成を模倣する視点、あるいは化学と酵素を組み合わせるchemoenzymatic synthesisが前に出ています。
2026年のJACSで報告されたスケーラブルなchemoenzymatic synthesisは、その象徴です。
ここで意味を持つのは、詳細手順の派手さではなく、TTXのような高機能天然物を、供給可能性を意識した形で再構成できる段階に入ったということです。
研究用試薬、構造活性相関、類縁体の比較、生合成経路の検証まで、一つの合成戦略が波及する範囲が広がっています。
ただし、課題もはっきりしています。
TTXは「作れる」だけでは足りず、天然由来の多様な類縁体群をどこまで系統的に扱えるかが次の壁になります。
実際の生物試料ではTTX単独ではなく複数の関連化合物が共存し、それぞれで毒性や分析応答がずれることがあります。
したがって合成研究は、天然物化学、分析化学、薬理学のいずれか一つに閉じるテーマではありません。
標準品供給を支える合成、検出法を支える分析、機能差を読む電気生理が一体化して、ようやくTTX研究の解像度が上がります。
2025年以降の総説を読むと、TTX研究はすでに「希少な天然毒の物語」から離れています。
現在の中心課題は、どう検出し、どう機能を測り、どう安定に供給するかです。
検出法の進歩と合成研究の前進が同じ時期に加速しているのは偶然ではありません。
測る技術が精密になるほど標準物質が要り、標準物質が増えるほど比較研究が進み、比較研究が進むほど生合成や毒性進化の仮説が絞られます。
TTXは今もなお、毒であると同時に、研究を前へ押し出す基準分子であり続けています。
フグが自分の毒で死なない理由は、毒を持っているからではなく、外から得たTTXを体内で蓄積・運搬しつつ、Na+チャネル側は耐性変異で結合しにくくなったという二層構造にあります。
毒は卵巣や肝臓に偏って集まり、産卵期の卵や仔魚の防御という生態的利点と結びついています。
ここを見ると、フグ毒は危険物質であるだけでなく、神経科学の基礎を切り開いた chemical tool でもあったとわかります。
記憶しておきたい芯はひとつで、フグが自前で毒を作ると単純化するのではなく、外因的なTTXの取り込み・体内動態・耐性変異を合わせて理解するということです。
(注)本サイトは現在コンテンツを拡充中で、毒物図鑑や関連解説記事を順次公開する予定です。公開後に本文中に内部リンクを追加して、関連記事へ案内します。
薬学部で毒性学を専攻し、製薬企業の安全性研究部門で毒性試験に従事。「毒と薬は紙一重」をモットーに、毒物の作用機序から医薬品への転換まで、分子レベルの科学をわかりやすく解説します。
関連記事
毒ヘビの種類と毒の違い|神経毒と出血毒を比較
「コブラ科は神経毒、クサリヘビ科は出血毒」と覚えると入口はつかめますが、毒性学の論文を読み込むほど、その図式だけでは取りこぼすものが増えていきます。3FTx、PLA2、SVMPといった毒素群の一次論文を概観すると、ヘビ毒は単一の化合物ではなく、タンパク質・酵素・ポリペプチドが混ざり合った複合毒であり、
トリカブトとアコニチン|日本三大有毒植物の科学
春の山菜採りの季節になると、毎年のように名前があがるトリカブトは、特定の一種ではなくAconitum属の総称で、日本ではドクゼリドクウツギと並ぶ「三大有毒植物」として記憶されています。
毒の進化|防御と捕食、盗用と共進化の科学
自然史系の特別展「毒」を歩くと、読者の頭にまず浮かぶのは「どれがいちばん危険か」より、「生きものはなぜ、こんな仕組みを持ったのか」という問いだと思います。この記事は、その疑問に答えるために、まずpoisonを食べたり触れたりして効く毒、venomを牙や針で注入する毒、
警告色と擬態|毒を持つ生物の進化戦略
林縁でスズメバチと、それによく似たハナアブを見比べていると、派手な黄黒模様は「きれいな色」ではなく、捕食者に向けた通信なのだと実感します。警告色は、有害性という二次防御を見やすい信号で正直に伝える戦略であり、その効果は捕食者が学び、似たパターンへ一般化できてこそ立ち上がります。